| Ойкологические деятели и их значение | Е. Варминг |
Свет оказывает влияние: 1) на питание. В отсутствии света нет ассимиляции угольной кислоты, нет жизни на земле. Начиная с известнаго минимума (различнаго для разных видов растений) ассимиляция увеличивается с возрастанием напряженности света, до известнаго максимума. Слишком сильное освещение влияет вредно; 2) на испарение, т. к. часть световых лучей превращается в растении в теплоту, содействующую испарению. В этом отношении существует также для каждаго вида растений известный оптимум. От слишкам сильнаго испарения растения защищают себя различным образом. Свет кроме того, оказывает влияние на явления роста, движения и вообще на все почти жизненные процессы.
Значение света для распределения растений. Нет почти такого уголка на земной поверхности, где-бы, благодаря недостатку освещения, растительная жизнь была невозможна, так как, если освещение слишком слабо в известное время года (напр., во время полярных ночей), то за то в другое время оно приобретает достаточную силу, что бы вызвать проявление жизни. Связанная со светом жизнь прекращается однако скоро по мере углубления в землю или в воду; на значительных глубинах могут жить только немногие, самые простые организмы.
Сила света имеет большое влияние на распределение видов и на богатство растительнаго сообщества неделимыми. В случае недостаточнаго освещения растения растут плохо, истощаются и гибнут. Общеизвестна разница между растениями, обитающими в лесах, тенистых местах, и растущими в местах освещенных. В полярных странах различие в облачности (числе солнечных дней, и дней пасмурных и туманных) является несомненно причиною описываемаго многими путешественниками различия между богатой флорой внутри фиордов и скудной растительностью побережий и островов области шхер (ср. Nathorst о Шпицбергене, Harty о восточной Гренландии).
О влиянии окраски и напряженности света на распространение водяных растений на разных глубинах будет речь в главе о гидрофитных растениях.
Время, в которое растение достигает своего полнаго разватия, находится в зависимости не только от силы света, но и от продолжительности освещения. Таким образом, если в Финляндии и Северной Норвегии ячмень созревает через 89 дней после посева, а в Шонене, не смотря наболее высокую температуру и большую силу света, для совершения той же работы нужно целых 100 дней, то причина этого заключается отчасти и в том, что более продолжительное освещение ускоряет образование вещества. На севере, благодаря более продолжительному освещению, периодическия жизненныя явления растений совершаются летом гораздо быстрее, нежели весной. По Арнеллю, чтобы зацвели растения в местности на один градус севернее Шонена в апреле нужно 4,3 дня, в мае 2,3 дня, в июне 1,5 дня и в июле 0,5 дня.
Время распускания листьев и цветков находится в зависимости от силы света. Часто сторона дерева, обращенная к источнику света, покрывается листьями раньше, чем противоположная сторона; напр, у бразильских видов Ficus сторона северная уже покрывается листьями, тогда как южная безлистна (Warming VIII); в полярных странах дерновины Silene acaulis часто бывают украшены цветками со стороны, обращенной к югу, куда направлены и цветки, между тем как часть, обращенная к северу, бывает вполне лишена цветков (Rosenvinge, Stefansson).
Сила света и направление световыхь лучей оказывают весьма большое влияние на форму растений.
Действие слишком слабаго освещения сказывается не только в явлениях этиолирования, что чаще всего представляет уже болезненное состояние растения, но оно бывает заметно и на здоровых, нормальных неделимых. Лесныя деревья представляют этому многочисленные примеры.
Прежде всего продолжительность жизни отдельных ветвей находится отчасти в зависимости от силы света. Затенение, производимое молодыми веками, препятствует ассимиляционной деятельности листьев более старых ветвей, развитию на них почек и в конце концов лишает их жизни, после чего сухия ветви обламываются ветром или под влиянием собственной тяжести. Вследствие этого вреднаго влияния внутренния части кроны у деревьев и кустарников не имеют листьев. Свободно растущая сосна имеет коническую форму и сверху до низу покрыта зелеными ветвями, между тем как такая же сосна в леснойчаще, благодаря разнице в освещении, имеет лишь маленькую зеленую крону, внизу же она совсем лишена ветвей или же покрыта сухими, лишенными листьев сучьями. Свободно растущия лиственныя деревья напр., буки, дубы и др. имеют вполне яйцевидную крону, тогда как растущия в густых зарослях лишь маленькую крону с поднятыми вверх ветвями (ср. рисунки Vaupell III).
Далее, отношение растений к свету играет важную роль в состязании, которое ведут друг с другом деревья, образующия сообщества Лесныя деревья могут быть подразделены на деревья, требующия много света и переносящия лишь незначительное затенение (светолюбивыя деревья), и на довольствующияся наоборот незначителным освещением и могущия существовать в тени (теневыносливыя деревья). Причины этого различия нужно искать прежде всего в специфических особенностях хлорофилла, затем в различной архитектонике видов (в строении побегов, расположении и форме листьев). Распределив наши лесныя деревья сообразно с их потребностью в свете, проявляющейся в их состязании, когда они растут вместе, и ставя наиболее светолюбивыя вперед, мы получим приблизительно следующие ряды.
1) Лиственница, береза, осина, ольха.
2) Pinus silvestris, P. strobus, ясень, дуб, вяз, Acer Pseudoplatanus.
3) Pinus montana Mill, ель, липа, граб, бук, пихта.
Замечательно и биологически важно обстоятельство, что почти все деревья в молодости могут переносить большее затенение, чем в более позднем возрасте. Дальше следует заметить, что способность переносить затенение находится в известной зависимости от плодородности почвы.
Разница между светомобивыми и тенелюбивыми растениями. Светолюбивыя и тенелюбивыя (гелиофильныя и гелиофобныя) растения разнятся между собою значительно как по своей внешней форме, так и по внутреннему строению.
Сильное освещение замедляет рост побегов; поэтому то гелиофильныя растения часто короткочленисты и сжаты, гелиофобныя же наоборот длинночленисты; растения, составляющия лесной ковер, обыкновенно высоки, с длинным стеблем. Листья светолюбивых растений обыкновенно узки, мелки, линейной или сходной формы, между тем как тенелюбивыя растения в тех же условиях имеют большие, широкие листья. Листья Majanthemum bifolium, растения, произрастающаго обыкновенно в тени кустарников, достигают на солнце всего 1/3, своей обычной величины.
Согласно Martins, Bonnier, Flahault и Scheutler'y листья многих видов растений достигают большей величины в северных странах, чем в широтах более южных, что должно по-видимому приписать большей продолжительности слабаго освещения.
Листья светолюбивых растений часто складчаты (злаки, пальмы, Pandanus), или кудрявы и бугорчаты, между тем как листья теневых растений плоски и гладки. Многочисленные примеры этого дает нам растительность сухих и жарких стран Вест-Индии (ср. напр. Johow I).
Направление листев также различно. На листья оказывает влияние самая незначительная разница в освещении, по отношению к которому они принимают наиболее выгодное для себя положение. Листья светолюбивых растений часто торчать прямо вверх, направлены почти вертикально (напр. Lactuca Scariola в солнечных местах и другия т. наз. компасныя растения; Stahl IV), или они свешиваются вниз, что особенно часто бывает у молодых растений (Mangifera Jndica и другия тропическия растения); между тем как листья тенелюбивых растений всегда распростерты горизонтально, что легко наблюдать на двудольных растениях наших буковых лесов. На листья светолюбивых растений солнечные лучи падают под острым углом, и следовательно не могут произвести полнаго действия, тогда как в лесах ослабленный свет падает на листья тенелюбивых растений под прямым углом.
У двудольных светолюбивых растений часто наблюдается образование т. наз. листовой мозаики (Kerner) состоящее в том, что маленькие и болышие листья соприкасаются своими краями и используют всю освещаемую поверхность (Fagus, Tnentalis, Mercurialis, Тгара). Разница между гелиофильными и гелиофобными растениями выражена особенно резко среди растений с игольчатыми и линейными листьями, напр. Juniperus, Calluna. Гелиофильныя растения имеют листья приподнятые вверх или прижатые, листья гелиофобных растений торчат во все стороны; у первых постоянно остающееся вертикальное расположение, у вторых изменяющееся горизонтальное; такия пространственныя отношения приобретаются растениями в молодости, во время роста. Упомянем здесь еще о фотометрических движениях, наблюдаемых у многих растений под влиянием изменений в напряженности и в направлены света. Под влиянием сильнаго освещения листья принимают профильное положение, более слабое освещение вызывает расположение горизонтальное. (Cp. 4 отд., 2 гл).
Анатомическое строение листьев светолюбивых и тенелюбивых растений представляет немаловажныя отличия. Листья светолюбивых растений часто равносторонни, если они занимают вертикальное положение, листья же тенелюбивых растений всегда двусторонни (дорзивентральны Heinricher, I). Листья светолюбивых растеши снабжены высокой палисадной паренхимой, состоящей или из одного ряда вытянутых клеток, или из клеток, расположенных в несколько этажей, или же из тех и из других одновременно (рис. 1).

Палисадная ткань теневых растений всегда невысока, (стебли, бедные листьями или совсем лишенные листьев, имеют обыкновенно высокую палисадную ткань вокруг стебля); зато губчатая ткань достигает у гелиофобных растений более мощнаго развития. Листья типичных гелиофобных растений состоять всего из одного ряда клеток (Hymenophyllaceae). Листья гелиофильных растений имеют узкия, листья гелиофобных растений широкия межклеточныя пространства. В одном и том же виде растеши процесс дыхания и ассимиляции совершается с большей интенсивностью в светолюбивых листьях, чем в листьях теневых (Lamarliere A. Мауег).
Кожица (эпидермис) светолюбивых растений толста и обыкновенно не содержит хлорофилла (она всегда лишена хлорофила на верхней стороне листа, Stohr); иногда она преобразовывается путем поперечнаго деления клеток в многослойную водоносную ткань (Ficus elastica и др. тропическия растения); ея кутикула, или кутикулярные слои, бывает всегда утолщена (рис. 2).
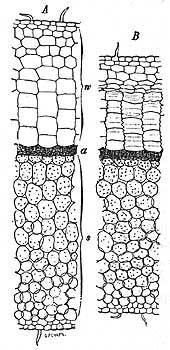
Кожица теневых растений тонка и однослойна, иногда содержит хлорофилл и покрыта тонкой кутикулой. Листья светолюбивых растений часто блестящи и отражают много света, чему представляют многочисленные примеры тропическия растения. Листья теневых растений матовы и завядают в сухом воздухе гораздо быстрее листьев светолюбивых растений. Эпидермическия клетки листьев светолюбивых растеши, в особенности на верхней стороне листа, имеют менее волнистыя стенки, чем у листьев теневых растений. Только нижняя поверхность двусторонних листьев светолюбивых растений снабжена устьицами или, по крайней мере, они здесь более многочисленны чем на верхней стороне (исключение представляют некоторыя альпийския растения) и погружены в ткань листа. У теневых растений устьица распределена равномерно на обеих сторонах листа, во всяком случае однако более многочисленны на нижней стороне, и вместе с тем лежат в одной плоскости со всей поверхностью листа или даже приподняты над нею.
Древеснение частей более распространено среди гелиофильных растений, напр. образование шипов. Благодаря отчасти этому обстоятельству, отчасти же большей толщине и строению кожицы листья гелиофильных растений обыкновенно жестки и кожисты; листья же гелиофобных растений по большей части тонки и, в случае большей величины, мягки (напр. листья многих из наших лесных растений, виды Corydalis и Circaea, Lactuca muralis, Oxalis Acetosella, многих папо-ротников, напр. в тропических странах Hymenophylaceae, мхов и пр.).
Степень волосистости весьма различна. Гелиофильныя растения, часто покрыты густыми волосками, серо-войлочны, серебристо-белы или иначе опушены, особенно на нижней поверхности (многия растения, растущия на скалах, на пустошах и в степях). Листья гелиофобных растений вообще гораздо менее волосисты, иногда даже совсем голы.
По всей вероятности существует большое различие в степени чувствительности хлорофилла к свету; хлорофилл гелиофобных растений, должно быть более чувствителен и обладает большей способностью использовать слабый свет, чем хлорофилл гелиофильных растений. Это подтверждается обстоятельством, что спиртовый экстракт хлорофилла папоротников весьма легко обезцвечивается на свету (Gautier).
Относительно влияния света на окраску растений нужно заметить, что помимо значения света для образования хлорофилла, он может еще, по-видимому, вызывать образование краснаго клеточнаго сока (антокиана). Под влиянием непосредственных солнечных лучей эпидермическия клетки голых частей растений окрашиваются нередко в красный цвет, что служит по-видимому защитой протоплазме и хлорофиллу (многие молодые побеги, проростки, высокогорныя и другия растения, хотя по Вилле окраска последних зависит м. б. и от влияния холода).
Кроме того, указывают (Bonnier, Flahault, Schuebeler и др.), что окраска листьев, цветков и плодов растенийв более высоких широтах более интенсивна, что, быть может, обусловливается действием почти непрерывнаго освещения.
Все эти вопросы будут разработаны более подробно в следующих отделах книги, особенно в четвертом (о ксерофильных растениях).
Из сказаннаго выше очевидно, что свет оказывает большое влияние на внешнюю форму и внутреннее строение растений. Это подтверждается еще способностью многих (м. б. большинства) растений приспособлять свое анатомическое строение и, главным образом, строение своих листьев к разным условиям освещения ("пластичные листья"). Лист бука напр., имеет на солнце иное строение, чем лист того же бука в тени (Stahl). Расположение хлорофилльных зерен в клетке и связанный с этим цвет листьев находятся в зависимости от освещения (Stahl, Sachs); более сильное освещение вызывает менее интенсивную окраску, и обратно. Что же касается физиологическаго объяснения этих влияний света, то должно сознаться, что мы еще весьма далеки от понимания, как и почему они происходят. Одни полагают, что свет, в зависимости от степени своей интенсивности, сам по себе вызывает упомянутыя различья в строении хлорофилльной ткани, но они не в состоянии выяснить, каким именно образом действует свет (Stahl, Pick, Mer, Dufourn др.). Другие придерживаются мнения, что причину явления нужно искать в усиленном испарении, которое опять-таки находится в зависимости от напряженности света (Areschoug, Vesque и Viet, Kohl, Lesage); еще другие приписывают главное значение усиленной, благодаря более интенсивному освещению, ассимиляционной деятельности листьев (Wagner, Мег, также Eberdt). Haberlandt предложил свою гипотезу о зависимости строения от образования веществ и от направления путей, по которым происходить передвижение материалов. Хлорофилльная ткань согласно этой гипотезе построена по двум принципам: принципу наибольшей поверхности, что должно содействовать процессу ассимиляции и принципу удаления продуктов ассимиляции по кратчайшему пути. Гипотеза эта, быть может до некоторой степени верная, не разрешает однако вопроса, почему строение листа меняется в зависимости от напряженности света, а также, почему в некоторых случаях клетки палисадной ткани располагаются косо к поверхности листа.
Почти не подлежит сомнению, что во всех вышеуказанных различиях между гелиофильными и гелиофобными растениями, мы имеем дело с проявлениями саморегулирующей способности растений (прямое приспособление, ср. отдел 7-й гл. 7-ю). Процесс саморегулирования совершается у нас на глазах в пластических растениях, способных приспособлять свое строение к степени напряженности света; в других случаях строение растения изменилось постепенно во время филогенетическаго развития и закрепилось путем наследственности в продолжение многочисленных генераций. Пользу всех этих различий в строении мы должны искать в следующем: в защите хлорофилла от разрушающего влияния слишком сильнаго освещения (Wiesner I); в защите самой протоплазмы (разрушающее действие света на протоплазму сказывается между прочим в том, что свет убивает бактерии, представляет собою дезинфецирующее средство), в защите против слишком сильнаго испарения и, наконец, в регулировании ассимиляционных процессов. Если принять во внимание, что мощность палисадной ткани находится в зависимости не только от напряженности освещения, но, как показали опыты, и от силы испарения, а также ото всех тех деятелей, которые содействуя поглощению растением почвенной влаги, влияют на испарение (напр., от солей в почве, повреждения корней), то поневоле приходим к убеждению, что самая существенная причина всех этих различий в строении состоит в регулировании процессов иcnapeния; это подтверждается еще и тем обстоятельством, что мощность палисадной ткани увеличивается заметно у растений, растущих в сухом климате. Испарение усиливается под влиянием более сильнаго освещения, т. к. световые лучи превращаются в растении в теплоту; свет самый важный деятель в процессе испарения, и растение регулирует его действие в зависимости от степени его напряженности. Окончательное решение этого вопроса принадлежит будущему.
Относительно затронутых здесь вопросов сравни работы: Areshoug'a, Stahl'я Pick'a, Dufour'a Haberlandt'a,
Heinricher'a, Vesque'a, Viet'a, Mer'a, Lothelier, Johow'a, Nilsson'a, Ebert'a и др.