| Мезофитныя сообщества | Е. Варминг |
В виду того, что сухое время в большей части стран в то или другое время года все же бывает, что даже в тропических лесах области дождей, например, на Яве, может быть такое время дня (перед полуднем, прежде чем в 2 или 3 часа дня начнется дождь), когда воздух относительно сух и испарение черезчур сильно (Haberlandt), - в виду этого листья большинства деревьев имеют различные способы защиты от слишком сильнаго испарения. Отчасти поэтому, а отчасти благодаря большей продолжительности жизни листа (более года), строение листьев оказывается более разнообразным, чем у листьев лиственных лесов умеренных стран.
В вечнозеленых тропических лесах листопад и появление новых листьев наступают не так одновременно, как в умеренных странах; изменения окраски листьев при этом не бывает. Листва опадает чаще постепенно, по мере того, как стареет; тем не менее, однако, например, в средней Бразилии, листопад происходить преимущественно в июле, августе или сентябре. Целый год лес имеет более темнозеленую окраску, чем обыкновенно наши леса; правда, некоторые виды во время появления листвы имеют особенно яркую окраску (обыкновенно молодые листья буро-красные), но они теряются в массе других видов. Почечных чешуи обыкновенно нет.
Так как листья деятельны, повидимому, в течение целаго года (некоторые виды почти весь год образуют новые листья), то не трудно понять, что растения способны добывать больше питательных веществ, чем наши деревья с опадающей листвой; этим объясняется быстрый рост и исполинские размеры многих тропических деревьев. Типов мезофильных вечнозеленых лесов несколько: во-первых, три собственно подтропических вечнозеленых типа, например, канарский лес из Lauraceae (который тем не менее, может быть, следует причислить к ксерофильным лесам), затем антарктические леса Южной Америки и тропические леса полосы дождей; кроме того, особые леса из определенных тропических растительных форм.
Класс сообществ: подтропические вечнозеленые лиственные леса.
Леса из Lauraceae Канарских островов описал Christ. В облачной полосе, где даже летом почти ежедневно бывают густые туманы, лавровые леса развиваются, главным образом, в долинах и ущельях. Почва покрыта густым зеленым ковром папоротников и мхов. Леса состоят из видов семейства лавровых (Persea Indica, Laurus Canariensis, Ocotea foetens, Phoebe Barbusana), к которым в большом количестве примешиваются Ilex Canariensis, Erica arborea, Myrica Faya и т. д. Подлесок состоит из Rhamnus glandulosa, Viburnum rigidum и др.; из лиан встречаются виды Smilax. Листья по большей части имеют форму лавровых, т. е. они нераздельны, цельнокрайни и кожисты; но есть и другие, чисто ксерофильные типы. В лесу под темным покровом лавровых деревьев господствует особая зеленая тень. Здесь всегда прохлада и влажность, составляющия резкую противоположность палящему зною открытых склонов, которыя усиливаются еще исходящим от лесной почвы запахом земли, мхов и фиалок. Почва покрыта почти исключительно подавляющей массой папоротников и напоминает леса на Новой Гвинее и других островах Тихаго океана; трав вообще встречается мало.
Класс сообществ: антарктические леса полосы дождей.
Антарктические леса стали известны из описаний Darwin'a и Hooker'a. Они распространены, начиная от южнаго Чили, от 36 град. южной широты, до Огненной земли, где они занимают пространство от моря до высоты в 1700-2000 m. на западном склоне цепи гор. Климат характеризуется невысокой годичной средней температурой (5-7 град.), отличающейся лишь на 9 град. от средней зимней и летней температур, но очень большим количеством дождей, распределенных почти по всем месяцам года. Благодаря этим условиям, развивается крайне богатый лес, который в северных своих частях массой лиан и эпифитов, а затем и подлеском, в котором важную роль играют древовидные папоротники и бамбук, производит впечатление тропическаго леса. По направлению к югу впечатление это теряется, но все же, вследствие сырого климата, деревья круглый год покрыты темно-зеленой листвой. Обыкновенным лесным деревом является здесь бук: Nothofagus antarctica, теряющий зимой листву, N. betuloides, N. procera, N. obliqua, N. Dombeyi, N. alpina и др. виды, все вечнозеленые. Листья этих буков малы, но многочисленны и напоминают миртовые; физиономия их поэтому совершенно другая, чем у нашего обыкновеннаго бука. Наоборот, лесная почва покрыта отчасти видами тех же родов, которые встречаются и в европейских лесах, например, Anemone, Viola, Sanicula, Galium, Vicia, Geranium и т. д. Кроме буков, значительную роль играют в лесу хвойныя (Araucaria, Libocedrus) и Proteaceae.
Флористически интересно то, что близкий к Fagus род Nothofagus растет здесь вместе с Proteaceae, Myrtaceae, Padocarpus, Libocedrus, Fitzroya Patagonica (хвойное дерево исполинских размеров) и другими тропическими и австралийскими типами и что на буке паразитирует Myzodendron. Особенно велико сходство с Новой Зеландией.
Класс сообществ: тропические леса полосы дождей.
Вокруг всего земного шара в экваториальных странах тянется лесная зона, которую всегда и подразумевают, когда говорят о первобытных лесах. Первобытным лесом называют, разумеется, всякий девственный лес, который сохранил свой первоначальный состав, причем не подвергался никакому или лишь незаметному влиянию человека, и деревья котораго стоят до тех пор, пока их жизнь не прекратится сама собой или в борьбе с соседями, пока мертвый ствол не упадет на землю, не истлеет и не оставит свободнаго места, которое скоро становится ареной борьбы для других видов. Еще несколько десятилетий тому назад первобытный лес существовал и в Европе; существует, разумеется, и теперь на "обвеваемой бурями скалистой почве Лапландии и Норвегии", также как в Германии (богемский лес) и на сырых и теплых равнинах Амазонки.
Тропические леса полосы дождей связаны с местностями, где дуют пассаты, где господствует высокая температура, где высоко стоящее на небе солнце посылает на землю потоки света и где вертикально поднимающияся, насыщенныя водяными парами массы воздуха, охлаждаясь и распространяясь в верхних слоях, вызывают ежедневно сильные ливни. Здесь между ветвями деревьев часто поднимаются теплые туманы, в известныя времена года большую часть дня с листьев дерев падают капли воды, и воздух может быть насыщен водяными парами (в Бейтенцорге на Яве влажность воздуха от 2-3 часов пополудни и до следующаго утра равняется почти 95%). Почва этого тропическаго леса всегда, разумеется, богатый гумус, черный и пористый, с гниющими остатками ветвей, листьев, цветов и плодов, взрываемый, конечно, разными животными. Тем не менее слой гумуса не так толст, как часто полагают; слой гумуса в несколько метров толщиной встречается далеко не часто (ср. Reinhardt и др.). Тогда как одни считают почву постоянно мокрой, другие и, разумеется, с большим основанием, говорят, что, благодаря пористости почвы, вода скоро просачивается.
При таких условиях растительный мир должен здесь развиваться с полнотой и разнообразием, какого нельзя найти больше нигде в свете. Даже если, как это доказывает Giltay (Ann. du jard.; de Buitenzorg, XV), образование веществ у тропических растений и не так велико, как это часто ошибочно говорится, все же тропический лес полосы дождей представляет высшую точку развития растительности на земле. Он имеет следующия особенности:
Использование места. Есть сообщества, вся площадь которых заселена очень тесно. Растения расположены во столько этажей, что все вместе они образуют сплошную путаницу ветвей. Получается "лес над лесом", как справедливо говорит Гумбольдт. Под верхним этажем деревьев, высокие, толстые, лишенные ветвей стволы которых подымаются иногда на высоту 40-50 m. растут другия деревья средней вышины, которыя не достигают ветвей перваго этажа, а под ним расположены и другие этажи: стройныя, тонкоствольныя, низкия пальмы, древовидные папоротники и др.; между ними кустарники из Urticaceae, Piperaceae, Myrsmaceae, Rubiaceae и др. Могучия травы, в 4-5 m. вы шиной, типа Scitamineae и Агасеае и другия травы примешиваются к кустарникам. Если остается свободное место почвы, куда может проникнуть свет, оно зарастает темно-зелеными папоротниками, Selaginella, мхами и тому подобными теневыми растениями (рис. 93). Но часто почва остается совершенно обнаженной, и черный гумус бывает покрыт только опавшими, гниющими и сырыми листьями, ветвями, остатками плодов и т. д., между которыми поселяются сапрофиты самаго страннаго вида (Burmanniaceae, Pirolaceae и т. д.), или же паразиты, живущие на корнях (Rafflesiaceae, Balanophoraceae) (рис. 93). Сюда же присоединяются массы эпифитов (рис. 7-12), которые покрывают стволы и ветви (Orchidaceae, Araceae, Bromeliaceae, Piperaceae и др. цветковыя растения, в Америке также и Cactaceae, затем папоротники, мхи и т. д.).

Деревья полосы туманов на Яве и Молуккских островах покрыты постоянно мокрым войлоком мхов, который бывает иногда толще самих стволов и придает им странный, темный вид. Из папоротников живут здесь, главным образом, мохообразныя Hymenqphyllaceae, которыя по своему анатомическому строению представляют "истинныя растения полосы туманов". Даже листья вечнозеленых видов могут быть одеты мелкими лишайниками и печеночными мхами. По мнению Schimper'a, всего более осадков требуют деревянистые эпифиты, из числа которых многие развиваются в лесах полосы дождей: огненно-красный Rhododendron Javanicum украшает кроны деревьев в горных лесах Явы и вместе с ним растут Ficus, Medinilla (Melastomaceae), Fagraea (Loganiaceae), Sciadophyllum (Araliaceae) и т. д. В яванских же горных лесах обыкновенны могучие папоротники Asplenium Nidus и Platycerium alcicorne, затем большие экземпляры Lycopodium Phlegmaria и др. видов Lycopodium и Psilotum (P. flaccidum), которые спускаются с деревьев, как длинные, в несколько метров длиной, лошадиные хвосты.

Наконец, мы видим здесь целую массу лиан, листья и цветы которых видны редко, за исключением опушек леса, где они спускаются каскадами цветов, но странные, длинные стволы которых висят тем не менее между почвой и вершиной деревьев, спускаются с них дугообразно или же стелются по земле.Многия другия растения дают лианам опору и помогают им достигать вершины деревьев. Причина такого богатаго развитая растительной жизни - освещение через редкия кроны верхняго этажа свет проникает к нижележащим кронам, а через них - еще глубже "Светлая полутьма" царит здесь, в таком лесу гораздо светлее, чем в нашем буковом. Все виды имеют, по выражению Юнгхуна, как будто "horror vacui" и с замечательным единодушием стремятся заполнить каждое пространство.
Но Tabulae Марциуса доказывают, что существуют леса, где среди исполинских колонн деревьев так темно, что не может развиваться почти никакая растительность.
Число видов тропическаго леса полосы дождей необыкновенно велико. Отсутствие совместной жизни индивидуумов одного вида, которое здесь наблюдается постоянно, стоит в полной противоположности с однообразием наших северно-европейских лесов и поясняется примером Бразилии, где на 3-х квадратных милях вокруг Лагоа-Санта в лесах растет около 400 различных древесных пород (Warming, VIII). Это разнообразие имеет несомненно геологическия причины, а именно - высокий возраст и никогда не прерывавшееся развитие тропической природы (Wallace, Warming, IX), отчасти физическия - выгодныя жизненныя условия, так как есть примеры, доказывающие, что сырая и плодородная почва производит большее количество видов, чем соседнря сухая и неплодородная.
Формы деревев. Большая часть форм не имеет ничего выдающагося, но некоторыя из них очень замечательны. Haberlandt (III) описал и изобразил некоторыя формы зонта, канделябра, многоэтажную и многия другия, уже известныя, не говоря уже о пальмообразных Ветвление несравненно разнообразнее и, по-видимому, много неправильнее, чем у наших деревьев; особенно часто бывает, что ветви несут пучки листьев только на своих концах и что каждый побег имеет мало боковых ветвей.
Досковидные корны встречаются у многих видов. Под этим разумеют корни, которые развиты много больше в вышину, чем в толщину, и которые отходят от нижней части ствола, часто на высоте 2-3 m, в виде больших, иногда изогнутых досок, поперечный разрез ствола представляет у самаго основания форму звезды со многими лучами и все пространство у подошвы дерева разделено на массу камер. Эти корни служат, разумеется, для того, чтобы деревьям с исполинским стволом и очень большой кроной дать твердую и широкую опору. Пластинчатые корни встречаются, главным образом, у Bombaceae и Ficus, далее у Mynstica, Carallia, Sterculia, Cananum и др. По Schimрег'у, они служат одним из главных признаков леса, получающаго обильные дожди, и отсутствуют в бедных дождями лесах.

Корни - подпорки других похожи на корни, встречающиеся, главным образом, у Rhizophora и имеются у некоторых пальм (Iriartea и др.), Pandanus. Они имеют вид цилиндрических подпорок, которыя отходят от ствола на известной высоте и спускаются к земле подострым углом, причем они имеют то же лучистое ветвление, как и у Rhizophora; число подпорок, которое имеет отдельный ствол, иногда весьма значительно (например более 20). Другую форму имеют они у Ficus religiosa и др., где они отходить от ветвей и дают каждому отдельному дереву возможность распространяться на огромную поверхность и образовать из одного дерева целый лес с необыкновенно густой и плотной кроной, которая дает глубокую тень; эта тень является, по-видимому, одной из причин столь сильнаго развития корней (рис. 94).
Колючие стволы нередки (Hura, Erythrma, Flacourtia и т. д.), чаще всего у пальм. Далее, здесь встречаются деревья с замечательными слоистыми корковыми наростами на стволах (Xanthoxylum и др.).
Почки не имеют (или имеют редко и в более сухих лесах) таких сухих почечных чешуек, как большая часть наших деревьев (Warming, VIII), но почки защищены травянистыми прилистниками, листовыми влагалищами, черешковыми выростами, а между почкой и ея покровом часто выделяется вода, смола или слизистая жидкость (Percy Groom).
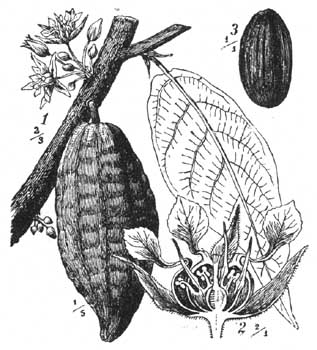
Цветков здесь видно немного, даже иногда поразительно мало; хотя тропический лес всегда очень богат цветками, но они всегда появляются слишком высоко, в кронах деревьев. Поэтому, если смотреть на лес с высокаго места, то часто можно видеть большия желтыя, белыя, фиолетовыя или красныя пятна: это цветущия деревья или лианы. Во многих случаях цветки очень мелки (например, у Lauraceae и у большей части Papilionaceae), но большое количество делает их заметными для насекомых. У некоторых видов цветки сидят замечательным образом на толстых ветвях или даже стволах, причем из года в год они появляются из тех же самых "спящих глазков". Самый известный пример таких растений - какао (Theobroma cacao, рис. 95); другие примеры дают Myrtaceae, Sapotaceae, Leguminosae, Ficus Roxburghu, Crescentia Cujete, виды Swartzia и др. (Wallace, Haberlandt, Esser in Verb, naturh. Ver. Rheinl. Wesif,1887, Huth в Verb. bot. Ver. Brand, 1888). Wallace думает, что цветки таких видов приспособлены для опыления бабочками, которыя часто порхают в тихом лесу. Насколько это мнение справедливо, до сих пор не выяснено. Для Theobroma, например, судя по строению цветов, это кажется несправедливым; здесь действуют скорее другия насекомыя или же происходить самоопыление.
Периодичность. В тропическом лесу нет ни зимы, ни лета, ни весны, ни осени; периодичность развития, замечаемая в других сообществах, здесь почти неприметна или же совершенно отсутствует.
Некоторые виды развивают в продолжение всего года новую листву; если некоторые виды и имеют ясно выраженный период покоя или стоят короткое время совсем без листьев и имеют определенное короткое время для развития новой, часто буро-красной листвы, то они совершенно теряются в массе других деревьев, не имеющих периода покоя или имеющих его в другое время года. Хотя большинство видов и имеет, по всей вероятности, определенное время для цветения, тем не менеедля разных видов оно всегда различно. Лес поэтому богат цветами круглый год (как саванны Южной Америки). Следовательно, жизнь тропическаго леса не имеет периодичности. Листья в тропическом лесу полосы дождей остаются на дереве почти всегда более года (обыкновенно около 13-14 месяцев; Warming, VIII) и сохраняют способность к деятельности часто много месяцев, может быть, более года, что имеет для растений огромное экономическое значение и объясняет их исполинский рост и производимое ими огромное количество органическаго вещества Старые листья, по Haberlandt'y, часто отгибаются, благодаря существованию активных движений, чтобы дать место молодым. Относительно окраски леса ср. приведенное на стр. 444.
Число форм листьев в тропическом лесу необычайно велико. Мы находим здесь не только встречающиеся у нас яйцевидные, эллиптические и тому подобные, простые или односложные листья, но и много других новых форм, например, перистую и веерообразную формы листьев пальм, большие, нераздельные, с особой нервацией листья Scitammeae, перистые листья Legummosae, особенно многосложный лист мимоз, безчисленные листочки которых производят движения, зависящия от силы света, пальчатые листья Bombaсеае и Panax (Arahaceae), пальчато-надрезные, щитовидные листья Cecropia и других, длинночерешковые, большие, сердцевидные или сердцевидно-яйцевидные листья Агасеае, затем злаковидные листья бамбука и т. д. Тем не менее самой частой формой является форма "лавроваго листа", т. е. большой, гладкий, блестящий, "эллиптический или ланцетовидный лист, примером котораго может служить лист Ficus elastica. Блестящие и кожистые листья - наиболее бросающаяся в глаза особенность тропическаго леса ("глянцевитость тропической листвы"), в то время как листья наших лесов матовы и прозрачны. Haberlandt говорить, что цельнокрайние листья здесь чаще, чем у нас. В общем легко заметить, что листья достигают часто исполинской величины, например, в сырых прибрежных лесах Бразилии и в лесах Амазонской реки (рис. 96); крометого, они много темнее, чем в умеренных странах, вследствие того, что листья и, главным образом, палиссадная ткань много толще, чем у нас. Другие же листья, особенно листья нижних этажей, напротив, очень тонки, вследствие слабаго освещения и большой влажности воздуха, которая здесь господствует.

Регулирование содержания воды в растениях. По изысканиям Haberlandt'a и других, растения яванскаго леса полосы дождей, особенно в верхних этажах, подвержены таким крайним условиям, которыя в нашей европейской природе не встречаются нигде. Приблизительно от 6-7 часов утра температура все повышается до 1-2 часов; постоянно возрастает и сухость воздуха под влиянием прямых лучей солнца. В конце-концов, воздух имеет часто дефицит насыщения в 30%. Другой период начинается около 2-3 часов дня грозами и страшными ливнями; остальную часть дня воздух так насыщен влагой (93-95%), что всякое испарение становится невозможным. Две трети дня воздух, следовательно, почти насыщен водяными парами. От этих опасностей, которыя в течение дня угрожают растениям с двух противоположных сторон, особенно их ассимиляционному процессу, они защищаются, как будет видно дальше, различными способами.
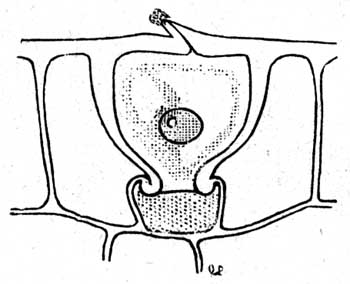
Если испарение задерживается упомянутым насыщением воздуха, то возникает опасность, что растения, вследствие постояннаго сильнаго давления корней, примут из мокрой земли слишком много воды, так что наступит "период высшей тургесценции", благодаря чему будет выдавлен воздух из межклетных пространств и они наполнятся водой. Эта опасность устраняется водоотделяющими органами, различные виды которых описаны "Haberlandt'м (V) под названием гидатод (т. е. водяных путей). Гидатодами мо гут служить: 1) клетки эпидермиса, иногда очень замечательной формы (рис. 97), или особые волоски (одноклетные или многоклетные, последние иногда в форме железистых волосков); так как эти органы помещаются на обеих сторонах листа, особенно снизу, то выделяющаяся вода имеет на листьях вид росы. 2) У некоторых палоротников гидатоды имеют вид железистых пятен на листовой пластинке. 3) Известныя водяныя поры, построенныя как большия устьица, часто встречающияся на верхней поверхности листовых зубцов над мелкоклетной, тонкостенной, обыкновенно безцветной тканью (эпитемой), в которой оканчиваются нервы. Затем вода может быть выделена без содействия гидатод, прямо через эпидермис, который имеет на наружной стенке поры. Наконец, вода может выделяться без содействия живых клеток, например, у злаков, в противоположность вышеприведенным случаям, где живыя клетки являются необходимыми и активными органами.
Другая опасность появляется благодаря сильной сухости воздуха и связаннаго с ней сильнаго испарения перед полуднем. По Haberlandt'y, в среднем испарение, правда, очень невелико, даже в 2-3 раза меньше, чем у растений средне-европейскаго климата (что, однако, Stahl (VI), Wiesner, Bourgerstein и Giltay оспаривают), но перед полуднем оно все же очень сильно и влечет опасность завядания или, во всяком случае, такого уменьшения тургора в растениях, что может потерпеть ассимиляция углекислоты. Этим объясняется то замечательное обстоятельство, что многия растения тропической полосы дождей имеют те же приспособления против сильнаго испарения, какия мы видели у ксерофитов: здесь мы находим толстый, сильно кутинизованный эпидермис, погруженныя устьица, слизистыя клетки, сосудистыя трахеиды, водяную ткань и т. п. Водяная ткань Ficus elastica хорошо известна. Давно известно также, что многия пальмы и большие и тонкие листья Scitamineae имеют на верхней поверхности или на обеих водяную ткань; она может быть так же сильно развита, как и ассимиляционная (Pfitzer, О. G. Petersen in Danske Vidensk. Selsk. Skrifter 6. R., Bd. VII, 1893); теперь эта водяная ткань становится понятной. Многие виды яванскаго тропическаго леса полосы дождей (например, Gonocaryum piriforme, Anamirta Cocculus) имют, по Haberlandt'y, в хлорофиллоносной ткани механическия клетки, совершенно как многие упомянутые на стр. 273 ксерофиты; оне имеют, очевидно, одну и ту же цель, т. е. защиту хлорофиллоносной ткани от сжатия во время засухи. Haberlandt нашел посредством опытов, что многия из описанных гидатод имели способность вбирать в себя растворы красящих веществ, и заключил отсюда, что оне служат и для принятия воды, и для проведения ея в растете. Это может происходить, разумеется, только в известные часы дня, а именно, когда проходят первые ливни, спустя несколько часов после полудня; если растение слишком сильно испаряло, гидатоды могут ему помочь быстро возстановить тургор листьев. Гидатоды являются, таким образом, регуляторами водоснабжения растений, которые то удаляют излишнюю воду, то всасывают ее, если имеется неотложная в этом необходимость. Все приведенное выше касается, разумеется, растений верхних этажей, листья которых находятся на поверхности леса и подвергаются действию солнечных лучей. У нижележащих растений, которыя прячутся в глубине леса и в тени, должны быть, напротив, другие признаки. Здесь мы, действительно, находим растения, которыя сильно приспособлены к тени и сырому воздуху, как, например, Hyme nophyllaceae, имеющия тонкие, как бумага, и немногослойные листья, без особаго эпидермиса и межклетных про-странств, стебли которых покрыты корневыми волосками и т. д. (Варминг: "Учебник ботаники", стр. 375).
Другия особенности строения находятся, как кажется, в тесной связи с тропическими ливнями, частью с их силой, не имеющей в нашем климате ничего себе подобнаго, частью с их изобилием. Приспособлена к механическому влиянию дождей. Шум дождя, падающаго на лес, слышен обыкновенно издалека и доказывает его силу, но деревья приспособлены к перенесению таких ливней, хотя они и способны вызывать сильное сотрясение, что отрицает, однако, Визнер. Многие простые листья очень тверды и кожисты; эпидермис может быть даже так сильно пропитан известью, что вся пластинка становится хрупкой и ломкой; она имеет вид покрытой "зеленым лаком жести". Листья других видов, особенно листья мимоз, акаций, других Leguminosae и пальм, разделены на массу листочков или долей, вследствие чего они представляют менее сопротивления падающему дождю; очень часто они имеют, сверх того, способность делать движения и свертывать свои листочки, подставляя каплям дождя наименьшую поверхность или даже один край. У других растений листья сложены складками или желобками, что делает их прочнее; это особенно ясно заметно у листьев пальм, листовыя доли которых расположены перисто или веерообразно, причем углы между складками обращены то вверх, то вниз. Черешки листьев часто направлены вверх; здесь, по-видимому, для другой цели, чем у ксерофитов, а именно для оказания ударам капель большаго сопротивления. Во многих других случаях листовыя пластинки и ветви висят вниз, особенно пока оне молоды, что имеет ту же самую цель; многие большие листья Araceae остаются на всю жизнь в таком положении, другие впоследствии выпрямляются. Исполинские листья пальм, Scitamineae и др. имеют большия стеблеобемлющия пластинки, которыя служат для того, чтобы сообщать стволу и листу большую крепость.
Обилие ливней также может действовать на растения неблагоприятно, так как пластинки листьев становятся слишком мокрыми и тяжелыми. Вследствие этого, испарение становится невозможным; эпифитныя водоросли, лишайники, грибы, печеночные мхи, даже бактерии (по Haberlandt'y) поселяются тогда на листьях и мешают ассимиляции углекислоты. Действительно, на более старых листьях многих вечнозеленых деревьев в сырых тропических лесах встречается масса эпифитных видов. Следовательно, для лесных растений полосы дождей, вероятно, выгодно, чтобы листья их высыхали скорее. По Jungner'y, Stahl'ю (V), которые относительно главнаго вопроса пришли к одинаковым заключениям почти одновременно, один в Камеруне, другой на Яве, быстрому высыханию способствуют различныя приспособления, а именно следующия: 1) Гладкая кутикула, которая не намокает; это очень распространенный способ. 2) Капельныя острия. Так называет Stahl длинныя заострения, которыми оканчиваются часто внезапно суживающияся пластинки и которыя типично развиты уFicus rebgiosa и др., но, кроме того, встречаются и у многих других растений (папоротников, однодольных и двудольных), как с простыми, так и со сложными листьями, и служат для того, чтобы отводить воду с легко смачиваемых листьев. Они направлены, разумеется, вниз, и чем длиннее острие, тем скорее сохнет лист. Саблеобразная форма заострения отводит воду всего лучше, иногда почти непрерывной струей (ср. рис. 3.) Капельныя острия никогда не встречаются у листьев с несмачиваемой поверхностью, а также у ксерофитов. 3) Часто встречаются также углубленныя жилки листа, которыя отводят воду к его кончику. Дугообразное жилкование листа у Melastomaceae и др. также удобно для этой цели. 4) Бархатистые листья встречаются, главным образом, у травянистых растений лесной почвы и у видов нижних этажей леса, где всего более тени и влаги. Клетки эпидермиса поднимаются в форме безчисленных низких сосочков, которые сообщают листу особый бархатистый блеск и между которыми вода, благодаря капиллярности, быстро разливается тонким слоем по всей листовой пластинке; этим достигается более легкое испарение воды.

Кроме того, относительно этих сосочков было высказано и другое мнение, именно, что они служат для лучшаго освещения внутренних частей листа (Stahl. VIII). Флора тропическаго леса полосы дождей так разнообразна, что описание каких-нибудь подробностей завело бы слишком далеко. Господствующая масса деревьев принадлежит к Leguminosae, Lauraceae, Myrtaceae, Moraceae и т. д. Леса, составленные из растений одного вида, в тропической лесной флоре - большая редкость. Леса тропической полосы дождей, вследствие большого смешения видов, составляют на всей земле, очевидно, только одно сообщество. Некоторые тропические леса появляются только при определенных условиях и представляют настолько отличныя жизненныя формы, что должны быть отнесены к отдельным сообществам: пальмовых, бамбуковых и папоротниковых лесов.
Класс сообществ: пальмовые леса.
К нижним этажам тропических лесов полосы дождей примешиваются также и пальмы, главным образом, невысокие, тонкоствольные виды. В тропической Южной Америке встречаются, однако, леса, где пальмы составляют подавляющее большинство видов; такие леса мы находим обыкновенно по берегам рек или на еще более влажной почве. В Бразилии встречаются, например, "Buritysales", т. e. леса из пальмы бурити (Mauritia vinifera и M. flexuosa). Lund пишет об этих лесах следующее: "Долины поросли свежим, ярким травяным ковром, а в глубине, где обыкновенно течет ручей, оне украшены группами несравненной по красоте бурити" (Warming, VIII); Martius в своих Tabulae изообразил леса из обоих видов. Затем большие пальмовые леса (из Copernicia cerifera) встречаются в северо-западной Аргентине, в равнинах Гран-Чако. Эти пальмы - световыя деревья, которыя образуют только редкие и лишенные тени леса; лес, таким образом, светел и имеет, вероятно, богатую травянистую флору (ср. также рис. 96).
Класс сообществ: бамбуковые леса.
Бамбуковый тростник (виды Bambusa) образуют в Восточной Азии почти непроницаемые леса. Гумбольдт говорит, что вдоль реки Магдалины растут непрерывные леса из бамбука и видов Heliconia с листьями вроде банановых. Тропическия реки часто окаймлены зарослями бамбука (ср. рис. 26).
Класс сообщества, папоротниковые леса.
В то время как пальмовые и бамбуковые леса требуют, по-видимому, довольно сырой почвы и должны, вследствие этого, правильнее всего причисляться к гидрофитным сообществам, древовидные папоротники связаны, главным образом, с влажностью воздуха; они составляют верный признак насыщеннаго водяными парами воздуха и равномернаго климата.

Леса Австралии и Тасмании богаты древовидными папоротниками. Вместе с другими папоротниками и тонколистными травами они образуют часто главную массу растительности. На многих более высоких Вест-Индских островах, например, на особенно богатой папоротниками Ямайке, на известной высоте в горах встречается растительность, которую можно назвать папоротниковым лесом (Cyathea, Alsophila), - может быть, слабое подобие одной из старейших флор земли (ср. рис. 99).